
Gli Anfibi
Sistematica
I Vertebrati sono suddivisibili in:
- Pesci, raggruppamento che riunisce i Vertebrati eterotermi che conducono vita acquatica e respirano attraverso le branchie.
- Tetrapodi, dal greco "tetra-" che significa "quattro" e "-podi" che significa "piedi", raggruppamento che riunisce i Vertebrati provvisti di arti e adattati all'ambiente subaereo (terrestre).
I Tetrapodi si distinguono in quattro classi:
- Anfibi
- Rettili
- Uccelli
- Mammiferi
Gli Anfibi rappresentano la classe che compie il passaggio dalla vita in ambiente acquatico alla vita in ambiente subaereo.
Durante il Devoniano superiore, più precisamente nel Famenniano (piano geologico compreso tra 374 e 359 milioni di anni fa) comparve il progenitore degli Anfibi, antenato comune a tutti i Tetrapodi, che abbandonò progressivamente l'acqua per portarsi sulla terraferma, già precedentemente colonizzata dagli Artropodi e dalle Piante.
Per tracciare i rapporti filogenetici che legano gli Anfibi ai pesci ossei occorre una breve digressione.
La classe degli Osteitti (pesci ossei) è distinta in due sottoclassi: gli Attinopterigi (nome che indica la presenza di pinne pari raggiate, si tratta dei pesci attualmente più diffusi e ricchi di specie) ed i Sarcopterigi, termine riferito alla pinna pari formata da fasci muscolari sostenuti da segmenti ossei, caratteristica che ha consentito la successiva evoluzione degli Anfibi. La grande radiazione dei Sarcopterigi risale al Devoniano, periodo geologico nel corso del quale divennero i pesci più diffusi. Attualmente sono scarsamente diffusi, in ridotta consistenza numerica e rappresentati da un esiguo numero di specie che vengono considerate "fossili viventi". Se ne distinguono due ordini: i Dipnoi ed i Crossopterigi.
La Latimeria, genere a cui è ascritto il Celacanto, è l'unico rappresentante vivente dei Crossopterigi. In origine anche i Crossopterigi, come gli attuali Dipnoi, erano provvisti di sacche polmonari, ma nella Latimeria tali strutture sono regredite poiché inutili nell'ambiente marino in cui essa si è adattata a vivere.

I Crossopterigi sono suddivisi in due gruppi: gli Attinisti, a cui è ascritta la Latimeria spp., e i Ripidisti, completamente estinti ma che hanno dato origine al progenitore degli Anfibi.

Gli attuali Dipnoi, come detto ascritti anch'essi ai Sarcopterigi, sono pesci dulciacquicoli distribuiti in prossimità della fascia equatoriale, il cui habitat è rappresentato da corsi d'acqua temporanei, soggetti a prolungati periodi di siccità causata dall'ingente evaporazione.
.jpg)
I Dipnoi si sono adattati a tali difficoltà ambientali evolvendo un polmone primitivo (perciò sono soprannominati "pesci polmonati"), originatosi dalla modificazione della vescica natatoria. Suddette condizioni ambientali sono simili a quelle avvenute su vasta scala durante il Devoniano superiore, perciò è plausibile che i progenitori degli Anfibi, che risalgono proprio a quel periodo, furono sottoposti alla medesima pressione selettiva degli attuali Dipnoi e che abbiano perciò evoluto modificazioni simili (fenomeno denominato convergenza evolutiva).

Indizi convincenti indicano che l'Osteolepis (estinto), il cui fossile fu rinvenuto nel nord della Scozia, faccia parte della linea evolutiva che congiunge i Crossopterigi Ripidisti agli Anfibi ancestrali.

L'Anfibio ancestrale più antico scoperto finora è l'Elginerpeton, il cui fossile fu rinvenuto in Scozia, nei depositi risalenti al Famenniano, precisamente datato 368 Ma (milioni di anni fa).

Un Anfibio ancestrale meglio conosciuto è l'Ichthyostega, il cui fossile fu rinvenuto in Groenlandia, negli strati risalenti al Famenniano, datato esattamente 363 Ma. Il corpo misurava 1,2 m di lunghezza ed aveva una forma simile a quella delle attuali Salamandre (Anfibi Urodeli, si veda più avanti) ma più tozzo.

L'Eryops è più recente rispetto all'Ichthyostega, essendo vissuto nel Permiano inferiore, circa 280 Ma. I suoi resti sono stati rinvenuti nel territorio attualmente occupato da vari stati meridionali del Nord America, tra cui il Texas. Il corpo era massiccio e dalle dimensioni insolitamente grandi per un Anfibio. Mostra inoltre maggiori adattamenti alla vita terrestre rispetto ai suoi predecessori.

Gli Anfibi primitivi si distinguono in due sottoclassi, entrambe completamente estinte:
- Lepospondili: il nome si riferisce alla particolare forma della vertebra, possedevano un corpo di piccola taglia ed ebbero minore successo.
- Labirintodonti: il nome si riferisce alla forma della corona dei denti (condivisa dai Crossopterigi e dai primi Rettili), possedevano un corpo di grossa taglia ed ebbero maggiore successo.
L'Ichthyostega è ascritto ai Labirintodonti, come anche i Rachitomi, gruppo che include l'Eryops; mentre il Diplocaulus era un rappresentate dei Lepospondili.

disegno di Dmitry Bogdanov.
Gli Anfibi primitivi raggiunsero il culmine della loro espansione durante il Carbonifero (periodo compreso tra 359 e 299 Ma). Il clima del Carbonifero era caldo-umido e le terre emerse erano spesso paludose e colonizzate da Artropodi che rappresentavano presumibilmente una fonte di cibo: una nicchia ecologica favorevole per gli Anfibi primitivi.
La grande estinzione permiana (225 Ma) coinvolse gravemente gli Anfibi primitivi causandone la scomparsa.

Gli Anfibi viventi sono denominati Lissanfibi per distinguerli dagli Anfibi primitivi estinti. I fossili testimoniano che i Lissanfibi, come li conosciamo oggi, in particolare Rane, Rospi e Salamandre, erano già presenti durante il Giurassico (compreso tra 199 e 145 Ma).
I Lissanfibi discendono da due linee evolutive distinte: gli Urodeli e Gimnofioni sono in rapporto filogenetico più stretto coi Lepospondili, mentre gli Anuri discendono direttamente dai Labirintodonti. Quindi tra Urodeli ed Apodi intercorre minore distanza filogenetica rispetto a quella che li separa dagli Anuri.
Pertanto Urodeli e Gimnofioni vengono raggruppati nella sottoclasse degli Urodeliformi a cui si contrappone la sottoclasse dei Batracomorfi che comprende gli Anuri.

Seguono gli aspetti classificativi dei tre ordini.
Anuri
Ordine a cui sono ascritti Rane, Ile (volgarmente dette Raganelle) e Rospi, il corpo è tozzo ed il capo non è visibilmente distinguibile dal collo, la coda regredisce durante lo stadio larvale ed è quindi assente nell'adulto, gli arti posteriori sono atti al salto e perciò più lunghi e robusti rispetto agli arti anteriori.

Urodeli
Anche chiamati Caudati, è l'ordine a cui sono ascritti Salamandre e Tritoni, il corpo è snello e provvisto di lunga coda, il capo è visibilmente distinguibile dal collo, gli arti anteriori e posteriori hanno pressoché la stessa dimensione, rappresenta l'ordine che meno sì è diversificato e morfologicamente più somigliante agli Anfibi primitivi.

Gimnofioni
Anche detti Apodi, ordine a cui sono ascritti i Cecilidi, il corpo è di forma cilindrica, estremamente allungato e privo di arti, risultato dell'adattamento alla vita ipogea (nel sottosuolo).

Con rare eccezioni, i Lissanfibi vivono nei sottoboschi umidi, al riparo dai raggi solari ed in prossimità di corsi d'acqua o di acque stagnanti poiché, malgrado adattati all'ambiente subaereo, non sono completamente emancipati dall'ambiente acquatico.
I Rospi sono gli Anfibi maggiormente adattati alla vita subaerea e perciò in grado di allontanarsi ad una discreta distanza dalle raccolte d'acqua, a cui però devono fare ritorno per la riproduzione.
Il vincolo degli Anfibi con l'acqua non risiede unicamente nelle caratteristiche dell'organismo adulto, ma dipende soprattutto dalle fasi del ciclo vitale quali accoppiamento, deposizione e stadio larvale. Gli Anfibi depongono le uova fecondate in acqua, e da esse si sviluppano i girini. In queste fasi del ciclo vitale l'ambiente acquatico è fondamentale, soltanto con la metamorfosi si avrà un individuo adulto capace di vivere sulla terraferma.
Come già spiegato nella dissertazione sulla Biologia dello Sviluppo, le uova degli Anfibi sono prive di guscio calcareo e soprattutto sono sprovviste di Amnios, annesso embrionale che svolge il ruolo di riserva d'acqua e che quindi consente la deposizione e lo sviluppo in ambiente subaereo. L'Amnios ha permesso a quei Tetrapodi che lo possiedono di emanciparsi completamente dall'ambiente acquatico e colonizzare le terre emerse.
La classificazione basata sulla biologia dello sviluppo suddivide i Vertebrati in:
- Amnioti (dotati di amnios), quali Rettili, Uccelli e Mammiferi.
- Anamni (privi di amnios), quali Ciclostomi, Condroitti, Osteitti ed Anfibi.
I Tetrapodi comprendono dunque sia Anamni (gli Anfibi) sia Amnioti (Rettili, Uccelli e Mammiferi).
La vita in ambiente subaereo ha comportato numerosi adattamenti che hanno interessato varie strutture dell'organismo, spiegati nel dettaglio nei seguenti paragrafi.
Tegumento
Come per gli altri Vertebrati, gli Anfibi presentano un tegumento composto da epidermide e derma. L'epidermide è a contatto con l'ambiente esterno, perciò l'epidermide dei Vertebrati a vita subaerea deve risolvere due problemi rispetto a quella dei Vertebrati a vita acquatica:
- impedire la disidratazione dovuta all'evaporazione,
- proteggere dalle radiazioni ultraviolette emesse dal Sole.
L'epidermide degli Anfibi deve contrastare la disidratazione non soltanto dei tessuti sottostanti ma anche delle stesse cellule di cui è composta, le cellule epidermiche degli Anfibi sono infatti cellule vive (al contrario dei Tetrapodi superiori nei quali l'epidermide è formata da cellule morte). L'adattamento si ottiene mediante l'acquisizione della cheratina, una proteina idrofobica che, prodotta e accumulata all'interno delle cellule epidermiche, crea una barriera contro la perdita di liquidi. Lo strato di cheratina viene periodicamente rinnovato. La cheratina è un'acquisizione che gli attuali Tetrapodi hanno ereditato dall'Anfibio ancestrale.
Nonostante la cheratina, le cellule epidermiche vive rappresentano un limite che impedisce agli Anfibi di colonizzare aree secche o poco umide.
Sebbene disporre di cellule epidermiche vive costituisca uno svantaggio per la ritenzione di liquidi, rappresenta un vantaggio riguardo agli scambi gassosi: gli Anfibi respirano in parte attraverso la cute. Oltre all'epidermide viva, gli scambi gassosi sono favoriti da un derma riccamente vascolarizzato. La scomparsa delle scaglie nei Lissanfibi (soltanto nei Cecilidi ne rimangano residui) è da attribuirsi alla necessità di scoprire l'epidermide per consentire gli scambi gassosi.
La radiazione ultravioletta è una porzione invisibile della luce solare che risulta dannosa per gli organismi viventi in quanto agente mutagene. Giacché la radiazione ultravioletta è riflessa entro un sottile strato d'acqua in prossimità della superficie, i Pesci non ne sono soggetti. La necessità di schermarsi dai raggi UV sorge dunque coi Tetrapodi. L'adattamento ai raggi UV si ottiene ricorrendo alla modifica funzionale di un pigmento preesistente (Preadattamento). I cromatofori sono cellule pigmentate, posizionate a diverse profondità sia nell'epidermide sia nel derma, comparse nei Pesci e la cui funzione originale è quella di colorare la livrea. Tra i diversi pigmenti alla base delle varie colorazioni, la melanina è in grado di schermare dai raggi UV, proprietà che viene sfruttata dai Tetrapodi, i cui principali cromatofori sono infatti i melanofori.
Differenti categorie di cromatofori hanno mantenuto la funzione originale conferendo, ad alcune specie di Lissanfibi, livree variopinte e sgargianti. Il colore giallo è dovuto agli xantofori, posizionati nella parte superficiale del derma, a contatto con l'epidermide; il colore blu è invece dovuto agli iridofori, posizionati più in profondità nel derma. Le varie colorazioni sono sotto il controllo dell'intermedina, ormone prodotto dal lobo intermedio dell'ipofisi.
Nella tela sottocutanea (lo strato più profondo del derma) degli Anuri sono presenti sacche linfatiche, borse ripiene di linfa, che garantiscono una certa riserva idrica. L'azione sinergica della cheratina e delle sacche linfatiche consente agli Anuri adulti, soprattutto ai Rospi, di allontanarsi dalle fonti di acqua più di ogni altro Anfibio.
Il tegumento degli Anfibi è talora provvisto di ghiandole pluricellulari (quindi con l'adenomero alloggiato nel derma ed il dotto escretore che si apre sulla superficie esterna dell'epidermide) a secrezione mucosa oppure a secrezione sierosa.
Le ghiandole a secrezione mucosa sono di grosse dimensioni e servono ad umidificare lo strato esterno dell'epidermide per impedire che si secchi a contatto con l'aria. Un altro tipo di ghiandole mucose si trovano nei cuscinetti cutanei, posizionati all'estremità delle dita di Hyla arborea, raganella in grado di arrampicarsi agilmente sugli alberi. Il muco prodotto da tali ghiandole ha funzione adesiva in quanto produce attrito a contatto col substrato. La seguente fotografia mostra una Hyla arborea posta su un vetro in modo da mettere in evidenza i cuscinetti cutanei.

Le ghiandole a secrezione sierosa sono le parotidi di alcuni Rospi e Salamandre, diverse dalle omonime "parotidi" dei Mammiferi: sono denominate entrambe "parotidi" perché posizionate dietro l'orecchio ("parotide" etimologicamente significa "presso l'orecchio"), ma mentre la parotide dei Mammiferi corrisponde alla ghiandola salivare, la parotide dei Lissanfibi produce un secreto irritante o velenoso di utilità difensiva. L'eventuale morso di un predatore (generalmente l'attacco si effettua sul capo della preda) rompe le ghiandole parotidi provocando il rilascio di tossine. Tale strategia difensiva risulta vantaggiosa sia per lo stesso individuo, che sopravvive all'attacco in quanto il predatore è dissuaso al primo morso, sia per la popolazione, poiché l'esperienza del predatore coinvolto funge da deterrente alla successiva predazione verso individui riconosciuti morfologicamente simili, spesso contraddistinti da colorazioni sgargianti, fenomeno denominato Aposematismo.
La taricatossina è una tossina molto potente prodotta dalle parotidi dei salamandridi del genere Taricha da cui prende il nome, viene utilizzata dagli indigeni dell'Amazzonia come veleno per i dardi.
Nel derma si trova uno strato mucopolisaccaride che conferisce al tegumento la capacità di reidratarsi per contatto con superfici umide. I Rospi dispongono di una placca cutanea posteriore specializzata, dotata di cute sottile e assorbente. L'acqua assorbita viene poi rilasciata quando necessario.
Durante il periodo di attività (per "periodo di attività" si intende il periodo in cui il metabolismo è regolare, in contrapposizione all'ibernazione e all'estivazione, si veda il paragrafo sul metabolismo), la pelle viene sostituita mensilmente. I brandelli di muta vengono asportati per mezzo degli arti anteriori ed ingeriti.
Per osservare un'eccellente fotografia istologica del tegumento dei Lissanfibi, si visiti la pagina Professor Udo Savalli (non posso caricarla sul mio blog perché i diritti sono riservati).
Scheletro
Lo scheletro degli Anfibi è osseo, proporzionalmente più massiccio di quello dei Pesci giacché i Tetrapodi, vivendo in ambiente subaereo, sono sottoposti ad una maggiore forza peso; di conseguenza anche la muscolatura si è irrobustita (la muscolatura si associa allo scheletro costituendo l'apparato locomotore).

Cranio
Il cranio è un neocranio protometamerico (per le definizione dei termini si consulti il compendio) e platibasico, quindi più primitivo rispetto al cranio dei Teleostei. Ciò non deve sorprendere in quanto gli attuali Teleostei sono più diversificati degli attuali Anfibi. Nel neurocranio platibasico (condiviso da Condroitti ed Anfibi) la piastra basale su cui alloggia l'encefalo è ampia. Tale caratteristica è contrapposta a quella del neurocranio tropibasico (dei Teleostei) nel quale lo sviluppo della piastra basale è condizionato dalla dimensione dei globi oculari: crescendo i due globi oculari si avvicinano medialmente comprimendo la base del cranio nella parte anteriore, di conseguenza l'encefalo arretra posteriormente rispetto agli occhi. Negli Anfibi la vista non è un senso particolarmente importante, per cui gli occhi sono di ridotte dimensioni e distanziati l'uno dall'altro, consentendo un neurocranio platibasico.
Gli Anfibi mostrano un tipo di sospensione dell'arcata orale innovativo rispetto ai Pesci, mentre in questi ultimi la sospensione è iostilica, gli Anfibi possiedono una sospensione autostilica costituita da un pezzo osseo di nuova acquisizione, denominato squamoso e compreso nello splancnocranio. Lo squamoso è presente nei Tetrapodi a partire dagli Anfibi. La necessità di sostituire la precedente sospensione con una nuova è dovuta all'assenza dell'iomandibolare, il quale si è modificato in columella, detta anche staffa. L'iomandibolare cambia forma, dimensione e si sposta all'interno del cranio arricchendo l'orecchio di una nuova componente, deputata al senso dell'udito. L'orecchio interno dei pesci ha funzione di organo statico, mentre la percezione delle onde sonore (in generale la percezione dei cambiamenti di pressione dovuti alla presenza di corpi nell'ambiente acquatico circostante) è assolta dalla linea laterale. Nei Tetrapodi la linea laterale scompare poiché non ha possibilità di impiego in ambiente aereo, e viene sostituita dall'aggiunta di una nuova camera dell'orecchio, detta orecchio medio. La columella fa parte dell'orecchio medio assieme alla membrana timpanica ed ai fonorecettori. La membrana timpanica si trova in corrispondenza dello spiracolo (canale che nei Condroitti serve a far passare l'acqua destinata alle branchie) ed è costituita da una cute sottile. La vibrazione che le onde sonore producono sulla membrana timpanica, viene trasmessa ai fonorecettori tramite la columella. I fonorecettori mostrano somiglianze strutturali coi neuromasti della linea laterale, dai quali presumibilmente derivano (per approfondimenti sui neuromasti, si rimanda al compendio). L'orecchio medio dei Lissanfibi è dotato di un unico ossicino, la columella, anziché tre come nei Tetrapodi superiori. L'orecchio medio serve principalmente agli Anuri per percepire le onde sonore ad alta frequenza dei richiami sessuali, prodotti dalle formazioni laringee e amplificati dalle sacche vocali, in occasione del periodo riproduttivo. Infatti negli Urodeli, che non adottano tali richiami, l'udito è poco sviluppato, e nei Gimnofioni è quasi assente. Le onde sonore di bassa frequenza sono invece percepite grazie ad un altro osso, l'osso opercolare (negli Osteitti l'opercolo serve a chiudere le branchie) che è associato ad un sottile muscolo opercolare, andando a formare l'apparato opercolare, situato in prossimità della columella. L'apparato opercolare consente di percepire rumori ambientali a bassa frequenza, come lo scorrere dell'acqua o il fruscio delle foglie.
Come nei Pesci, negli Anfibi è presente l'articolare dell'arcata inferiore ed il quadrato dell'arcata superiore, con la differenza che negli Anfibi il quadrato è articolato allo squamoso.
Lo splancnocranio degli Anfibi regredisce per via della scomparsa di tutte le arcate branchiali. Le branchie delle larve degli Anfibi sono esterne (come si vedrà meglio più avanti) e non associate al cestello branchiale come quelle dei Pesci. L'eliminazione delle arcate branchiali favorisce il sopravvento nelle proporzioni relative del neurocranio rispetto allo splancnocranio, tendenza che proseguirà nel corso dell'evoluzione dei Tetrapodi.
Nei Pesci le narici servono esclusivamente ad immettere acqua nella camera olfattoria (che contiene il tessuto olfattorio). Negli Anfibi invece ciascuna delle due camere olfattorie, oltre a sboccare all'esterno attraverso le narici, sbocca anche nella parte anteriore del palato, dietro i premascellari, attraverso due nuove aperture dette coane. L'aria introdotta ("inspirata" è inappropriato come si vedrà oltre) per la ventilazione entra dunque dalle narici e giunge ai polmoni attraversando le coane.
Mentre il cranio dei Pesci è fisso (vantaggio idrodinamico), quello degli Anfibi è mobile grazie ai condili occipitali, capi articolari posti sull'osso occipitale (regione posteriore del cranio) che collegano il cranio alla colonna vertebrale. I condili occipitali sono una nuova acquisizione dei Tetrapodi a partire dagli Anfibi. Il numero dei condili varia a seconda della Classe e viene utilizzato come carattere diagnostico per la determinazione dei reperti fossili. Sia gli Anfibi sia i Mammiferi possiedono due condili, condizione primitiva che non consente di ruotare il capo oltre pochi gradi. Gli Uccelli e i Rettili possiedono un unico condilo che permette un'ampia rotazione del capo. Un cranio capace di ruotare consente di scrutare rapidamente l'area circostante aumentando le probabilità di sopravvivenza.
Vertebre
La vertebra anficele ha la faccia cefalica concava e la faccia caudale convessa, viceversa la vertebra opistocele ha la faccia cefalica convessa e la faccia caudale concava. Gli Urodeli sono accomunati dalla vertebra opistocele, mentre negli Anuri la vertebra varia a seconda della specie e perciò rappresenta il carattere distintivo su cui si basa la classificazione dei sottordini, come si vedrà nel relativo paragrafo.
La colonna vertebrale dei Batracomorfi differisce dal quadro generale dei Tetrapodi. I Batracomorfi sono sprovvisti di gabbia toracica (indice di primitività), di conseguenza le vertebre toraciche sono indistinguibili dalle vertebre lombari: il processo trasverso ha la stessa lunghezza in entrambe. Pertanto nei Batracomorfi si parla propriamente di regione toraco-lombare e di vertebre toraco-lombari. Inoltre la coda è presente negli Urodelomorfi ma assente negli adulti dei Batracomorfi, perciò nella colonna vertebrale di questi ultimi sono riconoscibili soltanto tre regioni (anziché cinque come negli altri Tetrapodi). Gli Urodelomorfi, a differenza dei Batracomorfi, presentano coste, sebbene reciprocamente disgiunte.

Nei Mammiferi il numero di vertebre che compongono ciascuna regione è conservativo (si dice di carattere che rimane invariato nel corso dell'evoluzione): persino il topo e la giraffa condividono lo stesso numero di vertebre. Nei Lissanfibi invece, solo il numero di vertebre della regione cervicale e della regione sacrale è fisso. Tutti i Lissanfibi possiedono una singola vertebra cervicale ed una singola vertebra sacrale, carattere esclusivo nell'intero subphylum dei Vertebrati. Il numero delle vertebre delle restanti regioni (toraco-lombare ed eventualmente caudale) varia a seconda della specie. Nella maggior parte degli Anuri il numero di vertebre toraco-lombari ammonta a 7, sommando la vertebra sacrale e cervicale risulta un totale di 9. Alcune specie di Anfibi hanno un numero caratteristico di vertebre, tanto da fornire un criterio diagnostico per la determinazione dei reperti fossili. Il numero di vertebre dei Gimnofioni è molto più elevato, in alcune specie arriva a 300.
Arti
La locomozione dei Pesci dipende dalla propulsione della pinna caudale, nei Tetrapodi è dovuta agli arti, i quali derivano dalla modificazione delle pinne pari (che nei pesci servono a mantenere l'equilibrio) dei Crossopterigi Ripidisti. Dalla pinna dorsale deriva invece la cresta dei Tritoni. Prescindendo le diverse dimensioni, l'arto dei Tetrapodi è una struttura omologa alla pinna pari dei Crossopterigi Ripidisti.
La pinna dei Crossopterigi Ripidisti è detta monobasale dicotomica, il nome indica che un unico pezzo basale si collega da un lato al cinto e dall'altro a due pezzi radiali. I pezzi ossei sono chiaramente associati a fasci muscolari.


Allo stesso modo, l'arto dei Tetrapodi è formato da un pezzo osseo collegato da una parte al cinto e dall'altra a due pezzi ossei. Nel dettaglio:
- Stilopodio: primo segmento, si articola con il cinto, è sempre costituito da un singolo pezzo osseo che nell'arto anteriore prende il nome di omero, mentre nell'arto posteriore prende il nome di femore.
- Zeugopodio: segmento intermedio, si articola con lo Stilopodio, risulta sempre formato da due pezzi ossei che nell'arto anteriore prendono il nome di radio e ulna, mentre per l'arto posteriore prendono il nome di tibia e fibula.
- Autopodio: segmento finale, si articola con lo Zeugopodio, è composto da un maggior numero di ossa ed è perciò una struttura più complessa, è suddiviso in tre regioni:
- basipodio, detto carpo se anteriore e tarso se posteriore;
- metapodio, detto metacarpo se anteriore e metatarso se posteriore;
- acropodio, composto da falangi in relazione con il metapodio.

Il primo dito ha due falangi, dal secondo al quarto dito si hanno tre falangi, il quinto dito ha nuovamente due falangi (condizione più comune, sebbene alcuni Tetrapodi, tra cui l'uomo, hanno tre falangi anche nel quinto dito). Come mostra l'illustrazione, è anche presente una sorta di dito, composto da due pezzi ossei estremamente ridotti, tale struttura prende il nome di prepollice se si trova nell'arto anteriore, o prealluce se nell'arto posteriore. L'arto dei Tetrapodi è detto pentadattilo, termine che indica la presenza di cinque dita. Gli Anfibi primitivi erano invece esadattili, ovvero con sei dita, come dimostrano numerosi reperti fossili. Ulteriore conferma della discendenza dei Tetrapodi dagli Anfibi primitivi deriva dal fatto che sovente alcuni individui nascano con sei dita, specie nell'arto posteriore, e ciò accade sia nei Lissanfibi e più raramente anche nell'uomo. Fossili di Anfibi primitivi, rinvenuti alla fine del secolo scorso e tutt'oggi oggetto di studio, mostrano arti posteriori con sette e otto dita.
Nel corso dell'evoluzione dei Tetrapodi, zeugopodio e stilopodio rimarranno strutture conservative, mentre l'autopodio subirà notevoli modifiche.
Si suppone, senza inequivocabili conferme dei reperti fossili, che i primissimi discendenti dei Ripidisti evolsero un arto orizzontale, nel quale stilopodio, zeugopodio ed autopodio erano disposti sulla stessa retta e perpendicolari rispetto al cinto. La locomozione di questo tipo di arti è paragonabile al movimento dei remi di una barca, per cui la deambulazione doveva consistere in un lento strisciare per terra. I supposti Tetrapodi dotati di questo tipo di arti dovevano essere lenti ed incapaci di affrontare lunghi percorsi sulla terraferma. I reperti fossili mostrano chiaramente che gli Anfibi primitivi erano dotati di arti trasversali, i cui segmenti sono disposti a forma di "z": lo zeugopodio si è disposto verticalmente sollevando il corpo da terra. Quindi la distanza che separa la parte ventrale dell'animale dal terreno equivale alla lunghezza dello zeugopodio.

Gli arti sono collegati al tronco mediante il cinto (detto anche cingolo), distinto in cinto pettorale (anteriore) e cinto pelvico (posteriore), e saldamente ancorati grazie ad una robusta muscolatura.
L'arto di tipo trasversale è condiviso da Cheloni, Lacertiliani, Loricati (tutti ascritti ai Rettili) e dagli Anfibi Urodeli. Malgrado rappresenti un notevole miglioramento rispetto alla locomozione offerta dall'arto orizzontale, l'arto trasversale rimane affetto da un'imperfezione che limita la velocità dell'animale: la deambulazione si effettua spostando alternativamente in avanti l'arto destro e l'arto sinistro, movimento che tramite il cinto coinvolge la colonna vertebrale che, di conseguenza, si flette ritmicamente dalla parte opposta all'arto che viene spostato in avanti.

L'arto parasagittale è condiviso da Mammiferi ed Uccelli, il perfezionamento si compie grazie allo stilopodio che si piega formando una testa articolare che va ad inserirsi nella cavità del cinto. In questo modo l'arto non è più in rapporto diretto con la colonna vertebrale ed è indipendente dalle masse muscolari del tronco. Quando l'animale deambula il movimento viene interrotto dalla cavità del cinto e non si trasmette alla colonna vertebrale.
Ricapitolando, gli Urodeli hanno arti anteriori e posteriori uguali e di tipo trasversale, nei Gimnofioni gli arti sono regrediti, mentre gli arti posteriori degli Anuri sono atti al salto. Vediamo ora nello specifico quest'ultima tipologia di arto.
In fisica, dalla legge di Hooke consegue che tante più spire ha una molla, maggiore è la forza che esercita, principio sfruttato dall'arto posteriore degli Anuri, che si è modificato allungando i segmenti ossei di cui è composto, in modo da poter spiccare il salto.

Il bacino ha assunto una forma particolare: gli ilei si sono talmente allungati da diventare due bacchette disposte parallelamente alla colonna vertebrale. La regione caudale della colonna vertebrale è assente, ma sostituita da una protuberanza denominata urostilo, formatasi dalla fusione degli abbozzi delle vertebre caudali nel corso dello sviluppo embrionale. L'urostilo è provvisto di una cresta ossea verticale sulla quale si inserisce il capo muscolare, l'altro capo del muscolo è attaccato al femore.
All'arto si aggiunge un nuovo segmento costituito da due pezzi ossei, il tibiale ed il fibulare, che derivano dall'allungamento di componenti del tarso (autopodio). Anche le dita risultano notevolmente allungate. Oltre alla necessità di allungare i segmenti in modo da poter spiccare il salto, si consideri che questo arto è palmato, per cui le lunghe falangi servono anche ad armare la membrana.

Gli angoli che si vengono a formare tra i vari segmenti dell'arto posteriore in un Anuro accovacciato sono tutti angoli acuti. La meccanica del salto si compie allineando ciascun segmento dell'arto posteriore al piano dell'ileo: il femore ruota rispetto all'ileo di circa 160° e simultaneamente la tibia-fibula ruota rispetto al tibiale-fibulare.

Sistema Nervoso
Per la definizione dei termini utilizzati in questo paragrafo si consulti la parte propedeutica.
Nei Lissanfibi il tetto ottico è ancora il centro nervoso coordinatore.
Gli Anfibi sono animali dallo scarso dinamismo, di conseguenza il cervelletto è molto ridotto, meno sviluppato rispetto a quello dei Condroitti e degli Osteitti.
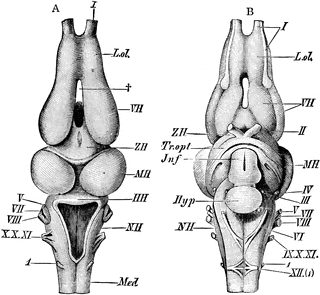
Come detto, negli Anfibi, all'orecchio interno (organo dell'equilibrio) preesistente nei pesci, si è aggiunto l'orecchio medio che è l'organo dell'udito. Corrispettivamente nel tegmento del mesencefalo si è sviluppato un centro nervoso che raccoglie le informazioni acustiche.
Nel telencefalo degli Anfibi, più marcatamente negli Anuri, si completa il processo d'inversione iniziato nei pesci cartilaginei e proseguito nei pesci ossei: le archicortecce superiormente e i nuclei del setto inferiormente, sono confluiti sul piano mediano giungendo a contatto. Pertanto è dagli Anfibi in poi che i Vertebrati mostrano due emisferi telencefalici perfettamente separati, ciascuno dei quali possiede il proprio ventricolo e la copia delle quattro aree.
Apparato Respiratorio
Le larve degli Lissanfibi vivono in acqua e sono provviste di branchie che verranno riassorbite nel corso della metamorfosi. Come precedentemente accennato, le branchie degli Anfibi sono analoghe a quelle dei pesci, ossia sono strutture di diversa derivazione.
La seguente fotografia mostra un esemplare di una specie neotenica, che conserva le branchie anche allo stadio adulto.

Le branchie degli Anfibi sono branchie esterne derivate dall'estroflessione dell'epitelio faringeo e riccamente vascolarizzate. Come si può osservare nella foto qui in alto, le branchie dipartono liberamente dalla regione ioidea e non sono incluse nel cestello branchiale (che non sussiste affatto). Questo tipo di branchie è condiviso dai girini sia degli Urodeli sia degli Anuri, sebbene in questi ultimi le branchie non siano visibili esternamente in quanto appena formatesi vengono ricoperte da una piega cutanea che funge da protezione e che lascia passare l'acqua attraverso un sifone; anche in questo caso si parla propriamente di "branchie esterne" poiché lo splancnocranio non è coinvolto.

Gli Anfibi adulti presentano polmoni sacciformi, si tratta di un tipo di polmone molto primitivo che consiste in un sacchetto al cui interno si abbozzano concamerazioni che offrono maggiore superficie all'epitelio respiratorio, il quale rimane comunque limitatamente esteso. Lo stesso tipo di polmone è condiviso dai Dipnoi, sebbene in questi ultimi risulti più concamerato di quello degli Anfibi. Il polmone deriva dalla modifica della vescica natatoria, organo che nei pesci ossei serve ai movimenti batimetrici, che a sua volta ha avuto origine da un'estroflessione dell'esofago.
La ventilazione polmonare dei Lissanfibi avviene in maniera diversa dagli altri Tetrapodi, in quanto i Lissanfibi mancano di gabbia toracica e di diaframma, né possiedono alcun muscolo che vicari la funzione del diaframma. Pertanto l'aria non può essere aspirata ma deve venire pompata. Per introdurre l'aria nel polmone i Lissanfibi mantengono la bocca serrata: l'aria che penetra nelle narici raggiunge il cavo orale attraversando il palato tramite le coane, perciò se la bocca fosse aperta l'aria fuoriuscirebbe anziché introdursi nei polmoni. I Lissanfibi insufflano l'aria nei polmoni per mezzo di movimenti del pavimento della bocca e della faringe (sostanzialmente l'aria viene "inghiottita"). Nel ciclo di ventilazione, diverse immissioni (analogo a "inspirazione") si alternano ad un'unica espirazione: il polmone ha una componente istologica elastica, le immissioni di aria si susseguono fino a dilatare il polmone al punto di provocare una contrazione che lo svuota. L'aria viene infine espulsa aprendo la bocca, perciò l'aria in uscita non attraversa mai le coane nella direzione opposta all'aria in entrata.

_dissection_-_(3).jpg)
A causa della scarsa efficienza dei polmoni, si rendono necessarie due ulteriori modalità di ossigenazione: la respirazione cutanea, già spiegata nel paragrafo relativo al tegumento, e la respirazione buccofaringea. Quest'ultima si effettuata mediante la mucosa del cavo orale che, venendo a contatto con l'aria destinata alle sacche polmonari, è capace di assimilare particelle di ossigeno.
Apparato Circolatorio
La seguente figura illustra molto bene l'anatomia comparata dei vari sistemi circolatori dei Vertebrati.

La circolazione degli Anfibi è doppia ed incompleta. Per "doppia" si intende che il sangue passa due volte dal cuore per ogni ciclo, "incompleta" significa che il sangue ricco di ossigeno si mescola in alcuni tratti con il sangue ricco di anidride carbonica.
Il cuore ha due atri separati, un unico ventricolo ed un unico cono arterioso dotato di un'unica valvola che crea una sorta di separazione. Il seno venoso si è notevolmente accorciato e, come nel caso dei Dipnoi, si è addossato all'atrio destro.

La duplicità del sistema respiratorio ha conseguenze sulla morfologia cardiaca. L'estensione dell'atrio destro è circa doppia rispetto al sinistro. L'asimmetria dipende dal fatto che nell'atrio destro confluisce sia il sangue refluo dei vari organi e tessuti (ricco di anidride carbonica), sia il sangue refluo dell'epidermide e dal cavo orale (ricco di ossigeno), mentre all'atrio sinistro giunge solamente il sangue ossigenatosi nelle sacche polmonari.
Dunque, diversamente da quanto accade nei Mammiferi (ove la circolazione è completa), nell'atrio destro confluisce sangue ossigenato mescolato a sangue ricco di anidride carbonica, mentre all'atrio sinistro giunge solo sangue ossigenato.
Dato che la sistole dell'atrio sinistro precede la sistole del destro, si crea una colonna di sangue composta in cima da sangue ossigenato, in basso da sangue ricco di anidride carbonica e nel mezzo si avrà un gradiente tra questi due poli. La ripartizione del sangue lungo suddetta colonna è importante poiché con la successiva sistole del ventricolo il sangue a diversa concentrazione di ossigeno verrà smistato in aree differenti del corpo. Spinta dalla sistole del ventricolo, la colonna di sangue attraversa il cono arterioso e la porzione ricca di ossigeno (posizionata in testa) verrà inviata alle carotidi giungendo all'encefalo, mentre la porzione mista (in posizione intermedia) verrà inviata all'aorta andando ad irrorare i vari organi e la parte periferica del corpo, ed infine la porzione povera d'ossigeno (posizionata in coda) verrà inviata all'arteria pneumo-cutanea che a sua volta si suddivide in arteria polmonare (che porta il sangue ai polmoni) e in arteria cutanea (che porta il sangue all'epidermide e al cavo orale).
Gli eritrociti degli Anfibi sono nucleati (carattere di primitività) e soffrono di una minore affinità per l'Ossigeno (lo catturano e legano con minore efficienza).
Sistema Digerente
I denti sono di dimensioni molto ridotte e di forma conica. La dentatura è di tipo omodonte, ossia tutti i denti hanno la stessa forma. Gli Anfibi sono polifiodonti, ovvero a ricambio continuo di denti, e l'impianto è pleurodonte, cioè il dente è semplicemente cementato sulla superficie dell'osso invece di essere alloggiato in una cavità. I Lissanfibi non masticano, i denti servono solamente a ghermire la preda per il tempo necessario ad ingurgitarla intera. I Rospi sono privi di denti.
La lingua muscolare compare nei Tetrapodi a partire dagli Anfibi, dato che nei pesci la lingua consiste in una bacchetta ossea (o cartilaginea nel caso dei Condroitti). La lingua muscolare è una struttura parecchio complessa, oltre ad essere costituita da fasci muscolari, è provvista di papille gustative ed innervata da tre tipi di nervi sensitivi ed un nervo motore che ne controlla il movimento.
Nella maggior parte degli Anuri ed in una minoranza di Urodeli, la lingua è infissa nella parte anteriore del cavo orale, anziché posteriore. In questi animali la lingua viene fulmineamente estroflessa per catturare la preda, che afferra grazie ad una superficie invischiante.
Tutti gli Anfibi adulti sono carnivori, predatori di Insetti, per cui l'intestino medio (tenue) risulta scarsamente convoluto: la digestione della carne, al contrario dei vegetali, non richiede tempi lunghi. Le ghiandole gastriche producono chitinasi, un enzima che idrolizza la cuticola chitinosa degli Insetti.
L’intestino si divide in intestino tenue ed intestino crasso e sbocca in cloaca, assieme ai dotti escretori e genitali.
_dissection_-_Stomach.jpg)
Sebbene gli adulti siano carnivori, i girini sono erbivori e si nutrono di alghe, pertanto l'intestino deve modificarsi durante la metamorfosi per adattarsi al cambiamento di dieta. La modifica si attua demolendo il precedente intestino e ricostruendone uno nuovo. Nel corso di tale rinnovamento l'animale non può alimentarsi, pertanto l'organismo ricorre all'assorbimento della coda per procurarsi i metaboliti necessari.
Apparato Escretore
Rene
Come tutti i Vertebrati, gli Anfibi hanno un prorene (detto anche pronefro) durante lo sviluppo embrionale, che viene sostituito da un mesorene (detto anche mesonefro) che, nel caso degli Anfibi, costituisce il rene definitivo (a differenza dei Tetrapodi superiori che lo sostituiscono ulteriormente con il metarene). Il tipo di mesorene degli Anfibi è molto simile a quello dei pesci dulciacquicoli. Sezionando la cavità addominale si osserva che il mesorene è molto esteso, addossato alla colonna vertebrale, e dotato di un canale, denominato dotto di Wolf, che trasporta i prodotti di scarto in cloaca. È da un'evaginazione del dotto di Wolf che nei Tetrapodi superiori avrà origine il metarene. Il mesorene, come il prorene, si forma dal peduncolo dei somiti. Per approfondimenti sullo sviluppo embrionale dei vari tipi di rene, si consulti il compendio.
Catabolismo
Lo smaltimento dei cataboliti azotati sotto forma di ammoniaca richiede un ridotto dispendio di energia ma un elevato consumo di acqua poiché, essendo eccessivamente tossica, l'ammoniaca deve essere abbondantemente diluita e frequentemente espulsa. Viceversa, lo smaltimento dei cataboliti azotati sotto forma di urea, richiede un più elevato dispendio di energia ma un limitato consumo di acqua. I Lissanfibi allo stadio larvale sono ammoniotelici, modalità privilegiata in quanto, vivendo in ambiente acquatico, dispongono di risorse illimitate di acqua. Allo stadio adulto diventano ureotelici, cambiamento imputabile alla necessità degli Anfibi terrestri di contenere la perdita di acqua. La vescica compare nei Tetrapodi a partire dagli Anfibi, ed è posseduta solo dai Tetrapodi ureotelici, giacché l'urea, non essendo tossica come l'ammoniaca, può essere raccolta e concentrata (con recupero di acqua). Nei Lissanfibi la vescica si collega ai lati della cloaca. Alcune specie arboree di Anuro sono uricoteliche, ossia smaltiscono i cataboliti azotati sotto forma di acido urico. La classe dei Lissanfibi comprende dunque tutte le possibili modalità di eliminazione dell'azoto.
Metabolismo
Gli Anfibi sono eterotermi, la temperatura corporea varia al variare della temperatura ambientale. Per mantenere il metabolismo a livelli ideali, i Lissanfibi rifuggono le aree esposte al sole, prediligendo i luoghi ombreggiati. L'eterotermia è un fattore che limita l'areale (ossia l'area occupata da un determinato taxon): i Lissanfibi sono assenti alle latitudini alte, ove il clima è rigido. I Lissanfibi distribuiti nelle zone climatiche caratterizzate da stagioni eccessivamente calde o fredde, si rintanano ed entrano in uno stato di quiescenza, che consiste nella sospensione delle attività e nella riduzione del metabolismo a livelli minimi. Durante la quiescenza, l'animale non si alimenta e l'organismo ricorre al consumo delle riserve di glicogeno accumulate durante il periodo di attività. Nelle zone temperate, durante la stagione fredda, i Lissanfibi vanno incontro ad ibernazione (o letargo); mentre nelle zone tropicali, durante la stagione calda, vanno incontro ad estivazione.
Anche durante il regolare periodo di attività, il metabolismo basale è moderato, in quanto, data l'eterotermia, non vengono sostenuti gli elevati consumi richiesti dal mantenimento della temperatura corporea stabile (omeotermia).
Ciclo Vitale
L'uovo è di tipo mesolecitico e la segmentazione è oloblastica diseguale, peculiarità dello sviluppo degli Anfibi è la semiluna grigia che si forma al lato opposto all'ingresso dello spermio.
L'uovo è dotato di un particolare involucro terziario (funzione protettiva) che prende il nome di involucro gelatinoso, costituito da glicoproteine che si idratano rapidamente a contatto con l'acqua e di conseguenza espandono il volume della struttura. Gli Anfibi depongono agglomerati di uova, detti ammassi, formati dalla reciproca coesione degli involucri gelatinosi. Tali ammassi possono essere informi oppure assumere la forma di cordoni o di calze o di fasce, a seconda della specie.



fotografie scattate da me e disponibili su Wikimedia.
È essenziale che la fecondazione esterna avvenga prima del contatto dell'uovo con l'acqua, giacché la struttura espansa dell'involucro gelatinoso rappresenta una barriera invalicabile per lo spermio, pertanto durante l'accoppiamento il maschio (di taglia più piccola rispetto alla femmina) si posiziona sul dorso delle femmine e rilascia gli spermi che colano sulle uova simultaneamente deposte dalle femmine. La deposizione delle uova nella femmina viene stimolata dalla stretta del maschio, in modo da coordinare le reciproche emissioni di gameti. Il maschio di Rana temporaria (riconoscibili dalla macchia posta nella regione temporale del capo) è munito di callosità del pollice per mezzo della quale stimola la femmina a deporre.

Generalmente gli Anfibi si avvistano proprio in occasione del periodo di accoppiamento, giacché escono allo scoperto per radunarsi presso le raccolte d'acqua.
Lo stadio larvale acquatico, detto girino, termina con la metamorfosi, mediante la quale si forma l'adulto adatto a vita terrestre. La metamorfosi è regolata dal rapporto fra concentrazioni ematiche di due tipi di ormoni: la maggior concentrazione di prolattina (prodotta dall'adenoipofisi) determina l'accrescimento larvale, quando la tiroxina (prodotta dalla tiroide) supera la concentrazione della prolattina si attiva la metamorfosi. Le rispettive concentrazioni di suddetti ormoni sono a loro volta sotto il controllo del TSH (ormone ipotalamico che stimola la produzione di tiroxina) e del PIH (ormone ipotalamico che inibisce la produzione di prolattina). È stato infatti osservato sull'Axolotl che la neotenia è dovuta alla mancata produzione di TSH.
Ecologia
I girini rappresentano la preda principale delle larve di Odonati (Libellule) e dei Ditiscidi (Coleotteri acquatici), gli adulti sono invece predati da Natrix natrix (Biscia dal collare), la quale ha sviluppato immunità alla tossina prodotta dalle parotidi.
I girini sono fitofagi, si nutrono di alghe. Gli adulti invece sono insettivori, si nutrono in particolare dell'imago degli Odonati: in questo caso preda e predatore coincidono (sebbene ciascuno nella rispettiva fase del ciclo vitale) e si instaura pertanto un controllo reciproco sulla crescita di entrambe le popolazioni.
Le Salamandre sono predatrici di Limacce (Molluschi).

Sottordini
Le specie ascritte agli Anuri vengono raggruppate in cinque sottordini:
- Anficeli
- Opistoceli
- Anomoceli
- Proceli
- Diplasioceli
I primi tre sottordini sono considerati arcaici giacché relitti. Un taxon (una qualsiasi unità tassonomica) è relitto nel caso in cui in passato abbia colonizzato una vasta area e successivamente, in seguito ad eventi sfavorevoli, l'espansione dell'areale si sia contratta lasciando popolazioni superstiti distribuite in zone circoscritte.

La classificazione dei sottordini degli Anuri è basata sulle differenze morfologiche fra le rispettive vertebre. Secondo alcuni autori le specie conosciute attualmente viventi ammontano a circa 3000.
Anficeli
Il sottordine degli Anficeli (il nome deriva dalla vertebra) conserva caratteri di primitività, tra cui principalmente le coste. Sono di piccola taglia (~5cm), il carattere distintivo è la pseudocoda muscolare utilizzata dal maschio come organo copulatore, per cui in questo sottordine la fecondazione è interna. Vengono raggruppati nella famiglia dei Leiopelmatidi che a sua volta comprende i generi:
- Leiopelmidi:
- in Nuova Zelanda; l'habitat è rappresentato da zone ad umidità satura ad elevate altitudini, privi di corsi d'acqua, per cui lo sviluppo larvale si completa nell'uovo da cui fuoriesce un piccolo morfologicamente uguale all'adulto, in altre parole manca il dimorfismo; una specie, la Leiopelma hochstetteri depone le uova all'interno dei cunicoli scavati dagli Odonati.
- Ascafidi:
- distribuiti negli USA Nord-occidentali; l'habitat è rappresentato da torrenti freddi e dalle correnti rapide a non meno di 2000 slm (metri dal livello del mare). Sono stenotermi (cioè tollerano intervalli di temperatura molto limitati) con soglia inferiore ai 10°C; il girino è provvisto di una ventosa cefalica per resistere alla corrente; grazie alla fecondazione interna possono accoppiarsi sul fondo dei torrenti; il periodo larvale è lungo, dura circa un anno, non si riproducono prima di sei anni dalla metamorfosi; mancano di membrana timpanica e di conseguenza sono muti; la colorazione è marrone variabile; pupille verticali; 9 vertebre presacrali.
Alcuni autori considerano suddetti generi come famiglie distinte.

Opistoceli
Il sottordine degli Opistoceli (il nome si riferisce alla vertebra) comprende le famiglie:
- Discoglossidi:
- diffusa in Eurasia con resti fossili nel Nord America; il nome si riferisce alla lingua di forma discoidale e aderente al palato, incapace di estroflettersi; suddivisa nei generi Discoglossus, Bombina, Alytes; al genere Bombina appartiene la specie Bombina pachypus, volgarmente conosciuta come "Ululone dal ventre giallo" è presente nelle aree collinari italiane a sud del Po; al genere Alytes appartiene la specie Alytes obstetricans, a vita terrestre, il cui maschio esibisce cure parentali verso le uova fecondate che trasporta sul dorso ed immerge periodicamente.


- Pipidi:
- detti Aglossi perché privi di lingua; vita permanentemente acquatica; dotati di linea laterale; arti posteriori ampiamente palmati; distribuiti in Sud America ed in Africa; la caratteristica peculiare riguarda le femmine che trattengono le uova fino a completo sviluppo (inclusa metamorfosi) inglobandole nella pelle del dorso.
.jpg)
- Rinofrinidi:
- famiglia costituita per un unica specie, la Rhinophrynus dorsalis, fossoria, vive all'interno di cunicoli che scava nel sottosuolo e che abbandona solo durante le piogge, in occasione delle quali si riproduce, le uova sono deposte in acqua e l'ammasso gelatinoso ha forma di tubo; il girino possiede lunghi barbigli; la metamorfosi dura pochi giorni e lo stadio adulto si raggiunge in pochi mesi; dimensioni estremamente ridotte (~3mm), colorazione marrone scuro, muso appiattito e di forma triangolare, sterno rudimentale, privo di denti, lingua non protrattile, si nutre di termiti e formiche; distribuito nella parte meridionale del Nord America e nell'America centrale (Texas, Messico, Giamaica, Yucatan).

Anomoceli
Le specie appartenenti al sottordine degli Anomoceli possiedono vertebre proceli (faccia cefalica concava), la vertebra sacrale è anchilosata con l’urostilo oppure ad esso articolata mediante un condilo. Al sottordine fa capo un'unica famiglia, quella dei Pelobatidi. Gli individui adulti di alcune specie raggiungono dimensioni ragguardevoli (~15cm). Comprende i generi Pelodytes, Pelobates e Scaphiopus variamente diffusi in Europa, Africa settentrionale, Asia minore, Asia sudorientale e Nord America. Sono fossori, vivono in zone aride perciò scavano allo scopo di rintanarsi nel terreno umido per mezzo di un tubercolo metatarsale a forma di mezzaluna; notturni; se spaventati emettono odore ripugnante come forma di difesa; si riproducono in pozze d'acqua temporanee perciò hanno uno sviluppo accelerato (dalla deposizione alla metamorfosi intercorrono solo due settimane); sussiste cannibalismo tra girini; pupille verticali; molte specie presentano una sporgenza tra gli occhi; ghiandola parotoide assente.

Proceli
Il sottordine dei Proceli prende il nome dell'omonima vertebra, inoltre gli individui appartenenti a questo sottordine presentano l'urostilo articolato alla vertebra sacrale mediante due condili. Ampia diffusione nelle regioni a clima temperato o tropicale di tutti i continenti. Comprende le seguenti famiglie.
- Leptodattilidi:
- alquanto estesa, conta circa cinquanta generi; forme eterogenee per taglia, habitat e abitudini (sia acquatiche sia terragnole e fossorie); distribuita in America centrale, Sud America e Australia; vi appartengono alcune specie dalle caratteristiche eccezionali, come Eleutherodactylus latrans adattatasi a vivere sulle formazioni calcaree delle regioni aride, è munita di una particolare ventosa posta sul ventre che le permette di aderire alle superfici rocciose, è inetta al nuoto, poiché l'ambiente non offre raccolte d'acqua, il maschio mantiene umide le uova bagnandole regolarmente con la propria urina; a questa famiglia appartiene anche il genere Ceratophrys conosciuto volgarmente come "rana cornuta" è tra i generi più allevati dai terrariofili, è originaria del Sud America ed è variopinta, robusta e molto vorace; il Telmatobius marmoratus è l'Anfibio che vive alla maggiore altitudine, è endemico del lago Titicaca (sulle Ande peruviane) a 3812 slm, per sopperire alla carenza di ossigeno dovuta alla rarefazione dell'aria ad alta quota, il T. marmoratus ha sviluppato evaginazioni cutanee per aumentare la superficie per gli scambi gassosi, inoltre gli eritrociti di questa specie esibiscono una maggiore affinità per l'ossigeno (riescono a catturarlo e legarlo con maggiore efficacia).

- Bufonidi:
- ovvero i Rospi propriamente detti, famiglia cosmopolita, vita terragnola, abitudini crepuscolari o notturne, legati all'ambiente acquatico solo per la riproduzione; forma tozza; cute verrucosa; scarso dinamismo dovuto alla protezione fornita dalle parotidi che gli conferisce un sapore che ripugna i predatori; la tossina secreta da Bufo alvarius viene utilizzata come stupefacente (effetti allucinogeni); sono estremamente prolifici, alcune specie arrivano a deporre fino a 35.000 uova; il genere Nectophrynoides del Madagascar ha sviluppato la viviparità per superare le difficoltà della siccità, la fecondazione è interna, l'accoppiamento si effettua congiungendo le reciproche cloache, la prole metamorfosa all'interno delle vie genitali della madre; un Rospo dall'aspetto insolitamente grazioso è il Bufo viridis, comunemente detto Rospo smeraldino, di piccole dimensioni, areale vastissimo (dall'Europa centrale fino alla Mongolia).

- Ilidi:
- conosciute come Raganelle; abitudini arboricole; zampe prensili dotate di lunghe falangi, all'estremità delle quali si trovano i bottoni adesivi; corpo snello e di piccola taglia; pupilla orizzontale; dotati di poderosa sacca golare grazie alla quale producono fragorosi richiami sessuali; famiglia cosmopolita, distribuita in America, Antille, Europa, Africa nordoccidentale, Indonesia orientale, Nuova Guinea, Australia, da livello del mare fino a 4500 slm; i generi Gastrotheca e Cryptobatrachus del Sud America annoverano specie le cui femmine trasportano la prole sul dorso fino a completamento della metamorfosi (cure parentali).

- Brachiocefalidi:
- capo di dimensioni cospicue, diffusi in America centrale e Sud America comprese le isole; nel genere Rhynoderma il cui maschio incuba le uova all'interno del sacco vocale, la prole fuoriesce dalla bocca terminata la metamorfosi; al genere Dendrobates appartengono gli Anuri dalle colorazioni più vivaci, aspetto dovuto all'aposematismo, sono infatti anche gli Anuri più tossici in assoluto, il nome del genere deriva appunto dall'inglese "dart-frog" (rana dei dardi) poiché vengono utilizzati dalle tribù indigene per ricavare il veleno per i dardi delle cerbottane da caccia (come per la Salamandra del genere Taricha); adottano cure parentali, il maschio trasporta la prole sul dorso, mentre alcune femmine depongono uova non fecondate per alimentare la prole; il genere Rhinoderma comprende un'unica specie, la Rhinoderma darwinii, descritta da Charles Darwin, di piccola taglia, molto dinamica, presenta una singola protuberanza sul muso simile ad un corno, le femmine depongono circa trenta uova che i maschi monitorano fino alla schiusa, momento in cui raccolgono i girini nell'ampia tasca golare e li mantengono fino alla metamorfosi.

- Microilidi:
- distribuiti lungo l'intera fascia tropicale di tutti i continenti; habitat caldo-umido; apertura orale stretta, specializzata nella cattura di formiche e termiti; comprende specie con abitudini arboricole, acquatiche e fossorie; il carattere distintivo della larva risiede nella mancanza dei denti cornei e della cartilagine della mascella, il labbro inferiore multipieghettato che consente una lunga estroflessione ad imbuto, uno spiracolo mediano; al genere Breviceps appartengono specie fossorie che hanno sviluppato protuberanze atte a scavare, simili a quelle dei Pelobates; arti molto corti e corpo sferoidale; vivono in zone aride e non entrano mai in contatto con l'ambiente acquatico, le larve si sviluppano all'interno dei cunicoli scavati dai genitori.

Diplasioceli
Il sottordine Diplasioceli è contraddistinto da colonna vertebrale che, a partire dal capo in direzione della coda, ha le prime sette vertebre proceliche, l’ottava anficelica e la nona è la sacrale il cui corpo vertebrale è convesso nella faccia cefalica e caudalmente si articola con l’urostilo mediante due condili. Si suddivide nelle due seguenti famiglie.
- Ranidi:
- ossia le Rane vere e proprie; reperti fossili dimostrano che questa famiglia esisteva già nel Miocene (da 23 a 5,3 Ma); si suppone che il centro di diffusione si sia trovato in Africa, continente ove tuttora è presente il maggior numero di generi; cosmopolita, assente solo in Groenlandia e Nuova Zelanda; forma idrodinamica, muso appuntito, vita sottile, arti lunghi e spesso palmati, di colore verde "Rane Verdi", pelle liscia, pupilla orizzontale, ampi timpani; vivaci; diurne; trovano impiego nell'alimentazione umana e anche impiego scientifico (a partire dai celebri esperimenti sulla "elettricità animale" di Luigi Galvani), fino a poco tempo fa utilizzati per i test di gravidanza (la gonadotropina delle urine delle donne gravide induce la produzione di uova nelle rane entro 48 ore); molto diffuse, il loro potenziale riproduttivo ne compensa il prelievo umano, depongono dalle 5.000 alle 10.000 uova in ammassi di circa 300 unità e raggiungono la maturità sessuale entro il primo anno di età.

- Racoforidi:
- l'arto presenta falangi intercalari cartilaginee; diffuse in Africa centrale e meridionale, Madagascar, Asia meridionale, Giappone, Filippine e Indonesia; colori brillanti, in particolare il genere Mantella del Madagascar; le specie africane entrano in estivazione durante la stagione secca e si attivano rapidamente al sopraggiungere delle piogge; alcune specie depongono le uova sulle foglie in prossimità degli specchi d'acqua, successivamente i girini anguilliformi strisciano lungo la pianta raggiungendo l'acqua; al genere Rachophorus del Borneo appartengono forme arboricole che utilizzano l’ampia palmatura degli arti per planare nell’aria spostandosi da un ramo all'altro, la specie più celebre è Rhacophorus reinwardti, volgarmente chiamata Rana volante, capace di planare per circa 15 metri grazie all'estesa superficie delle zampe palmate.
2.jpg)

Sono conosciute circa 400 specie viventi ascritte agli Urodeli, raggruppate in cinque sottordini:
- Criptobranchi
- Ambistomatoidei
- Salamandroidei
- Proteidei
- Meanti
Si tratta di animali schivi e legati al clima temperato, a differenza di Anuri e Gimnofioni non se ne segnala la presenza ai tropici. L'ordine annovera specie che conducono vita cavernicola, si rinvengono nei bui recessi delle grotte. Come già detto, conservano la coda allo stadio adulto. Sono privi di cavità timpanica e membrana timpanica. Gli Urodeli, in particolar modo le Salamandre, possiedono la prodigiosa capacità di rigenerare tessuti danneggiati, persino arti accidentalmente amputati. Tale capacità dipende da cellule totipotenti che permangono nell'adulto.
Criptobranchi
Il sottordine dei Criptobranchi è suddiviso in:
- Cryptobranchidae:
- ritenuta la famiglia più primitiva; completamente acquatici; descritte soltanto tre specie attualmente viventi, ascritte a due generi diversi, il genere Crytobranchus del Nord America e l'Andrias di Cina e Giappone; alla famiglia appartengono gli Anfibi di taglia più grande, l'Andrias japonicus è la salamandra gigante giapponese che raggiunge i 150cm di lunghezza, l'Andrias davidianus è la salamandra gigante cinese e misura 100cm, il Cryptobranchus alleganiensis è la salamandra alligatore americana e misura 75cm; i membri delle specie asiatiche trovano impiego nell'alimentazione umana; il corpo, compreso il capo, è appiattito e consente di incunearsi sotto le rocce; sebbene la metamorfosi sia parziale, gli adulti sono privi di branchie, gli scambi gassosi avvengono principalmente attraverso la cute coadiuvati dall'ondeggiamento del corpo; i maschi esibiscono cure parentali proteggendo le uova all'interno di nidi sommersi appositamente creati scavando sul fondo dei corsi d'acqua.

- Inobidae:
- distribuita in Russia, Asia paleartica e sud-orientale; a fecondazione esterna poiché le femmine sono sprovviste di ricettacoli cloacali per le spermatofore; polmoni ridotti, gli scambi respiratori si effettuano prevalentemente attraverso la respirazione cutanea e buccofaringea; il corpo misura dai 10 a 25 cm di lunghezza a seconda della specie; alcune specie sono neoteniche; il genere Onychodactylus di Cina e Giappone presenta artigli (carattere esclusivo).

Ambistomatidi
Il sottordine Ambistomatidi è endemico del Nord America. Ricco di specie simili per forma ed abitudini, si distinguono in base alla colorazione della livrea. Un osservatore inesperto potrebbe confonderli con le lucertole (Rettili), ma a differenza di queste non hanno pelle squamata, artigli, e canale timpanico. Spesso terragnoli, di abitudini notturne. Presentano ampio capo, occhi ridotti muniti di palpebre, pelle liscia, coda appiattita latero-ventralmente, più lunga nei maschi. Un carattere distintivo risiede nell'assenza della scanalatura nasolabiale. Durante il periodo riproduttivo, nei maschi sporge la cloaca. Le uova impiegano circa un mese a schiudersi, il periodo larvale si protrae per un anno. Annovera diverse specie neoteniche, i fenomeni di neotenia sembrano correlati alla scarsa concentrazione di iodio nei corsi d'acqua (condizione tipica del territorio occupato dagli stati del Wyoming e Colorado). A questo sottordine appartiene il celebre axolotl, forma neotenica il cui mutante albino è stato importato in tutto il Mondo per essere allevato in cattività; il nome scientifico è Ambystoma mexicanum, è originario del lago di Xochimilco che si trova nei pressi di Città del Messico; l'habitat dell'axolotl è rappresentato da acque ferme e ricche di vegetazione; la popolazione selvatica è vulnerabile e risente delle modifiche del territorio relative all'agricoltura; è predatore di vermi, molluschi e pesci; il periodo di accoppiamento varia da fine inverno ad inizio primavera, in seguito al corteggiamento il maschio rilascia una spermatofora che viene raccolta dalla femmina che la ripone all'interno della cloaca, quindi le uova vengono fecondate durante la deposizione venendo a contatto con la spermateca conservata nella cloaca femminile (stessa modalità dei Tritoni); gli ammassi di uova hanno forma di grappoli, ciascuno conta dalle 50 alle 100 unità, le larve si sviluppano in un mese; longevo, vive fino a 25 anni.

al sottordine è anche ascritto l'Ambystoma maculatum, l'habitat è rappresentato da boschi umidi decidui attraversati da corsi d'acqua; abitudini notturne; occhi ben sviluppati; dorsalmente la livrea è nero-bluastra cosparsa di macchie giallo o arancioni, sul ventre è grigia; arti corti, tronco robusto che reca dodici solchi costali visibili, la lunghezza varia dai 18 ai 25 cm.

Salamandroidei
Il carattere distintivo del sottordine dei Salamandroidei sono i denti posti sul palato, dietro le coane. La fecondazione è interna.
La famiglia Salamandridae ha vasta distribuzione, si trova lungo la fascia temperata di Europa, Asia e America. Alla famiglia appartiene il genere Triturus, il primo Urodelo descritto (da Linneo). Comprende le specie più note, i Tritoni (acquatici) e le Salamandre (terragnole). Alcune specie sono ovovivipare. Come già precedentemente accennato, le Salamandre producono secrezioni cutanee tossiche, pertanto presentano colorazioni aposematiche, spesso esibite assumendo una postura denominata unkenreflex: si ribaltano sul dorso per mostrare la colorazione ventrale di ammonimento all'osservatore. Molto particolare la specie Notophtalmus viridescens per via della doppia metamorfosi (dalla larva si sviluppa un adulto terragnolo il cui habitat è la lettiera dei boschi, dopo qualche anno metamorfosa ulteriormente diventando acquatico) e per il corteggiamento (il maschio afferra anteriormente la femmina per mezzo degli arti posteriori). Pachytriton è un genere endemico delle regioni montuose della Cina, zampe corte, la coda è a forma di spatola, estremamente compressa lateralmente e con la punta arrotondata, l'habitat è rappresentato dai torrenti freddi e poveri di risorse alimentari, per cui tale genere ha sviluppato un forte istinto territoriale ed aggressività intraspecifica; le femmine adottano cure parentali, depongono le uova nella tana (sotto le rocce) e le ventilano con movimenti della coda favorendone l'ossigenazione.

Il genere Salamandra è endemico dell'Europa; in Italia sono presenti la Salamandra salamandra (con due sottospecie la S.s.salamandra, autoctona delle Alpi, Prealpi e Appennini settentrionali e la S.s.gigliolii endemica degli Appennini del centro-sud), la Salamandra lanzai e la Salamandra atra; sul dorso la livrea è nero lucente costellata di macchie gialle, sul ventre è nero uniforme; la S.atra non ha macchie, viceversa nella sottospecie S.s.gigliolii l'estensione delle macchie supera lo sfondo nero; il dimorfismo sessuale è limitato al rigonfiamento della cloaca del maschio durante il periodo riproduttivo; l'habitat è rappresentato principalmente dai boschi decidui in zone collinari, ma si rinviene anche in boschi ad aghifoglie fino a 1800m e nella macchia mediterranea; abitudini crepuscolari e notturne; predatore di lombrichi, limacce e larve di varie specie di insetti; nonostante le secrezioni irritanti, è predato da Natrix natrix (biscia dal collare) che ha sviluppato immunità a tali tossine; il periodo di attività è in primavera ed autunno, mentre in estate ed inverno entrano rispettivamente in estivazione ed in ibernazione; il periodo riproduttivo è in primavera, l'accoppiamento avviene in ambiente subaereo, il maschio si posiziona sotto la femmina e depone una spermatofora sul terreno, dopodiché si sposta permettendo alla femmina di raccogliere la spermatofora all'interno della cloaca, la femmina trattiene le uova fecondate (ovoviviparità), ultimato lo sviluppo depone le larve in una raccolta d'acqua; insolitamente longevo per un Anfibo, vive fino a 50 anni; viene allevata in cattività dai terrariofili; la convenzione di Berna vieta il prelievo in natura di individui selvatici, per detenerle è obbligatorio possedere il certificato che ne attesta l'acquisto presso rivenditori autorizzati dunque la provenienza da allevamento.

fotografia di Iacopo Buttini.
Salamandrina terdigitata, soprannominata Salamandrina dagli occhiali (per via della macchia chiara sul capo), è un altro endemismo dell'Appennino italiano; diffusa soprattutto sul versante tirrenico, dalla provincia di Genova fino all'Aspromonte, l'habitat d'elezione sono i boschi di latifoglie tra i 200 e i 900 slm; è stato osservato che per la riproduzione usufruisce di piccole raccolte d'acqua artificiali, come i fontanili (questo la rende vulnerabile a causa degli agricoltori che svuotano i fontanili che contengono i girini); abitudine notturna, lucifuga (rifugge la luce), dimensioni estremamente ridotte (massimo 11cm le femmine e 9cm i maschi, ma la coda è i 2/3 della lunghezza), tutte caratteristiche che la rendono difficile da avvistare; carattere esclusivo della specie risiede nel possedere solo quattro dita, sia negli arti anteriori sia posteriori; tossicità e relativa colorazione aposematica ventrale esibita per unkenreflex; la disposizione dei colori del ventre, (bianco, rosso e nero) caratterizza ciascun individuo; oltre l'unkenreflex può eventualmente ricorrere alla tanatosi, ovvero si finge morta (l'etimologia deriva dal greco "tanatos" che significa "morte"); entrano in estivazione ma sono attivi d'inverno; i girini di questa specie vengono predati dai girini di Triturus carnifex e dai Ditischi (Insetti).

fotografia di Iacopo Buttini.
Il genere Triturus è diffuso in Eurasia; la specie Triturus vulgaris è la più adattabile e comune, corpo esile, livrea punteggiata; il Triturus helveticus, soprannominato "Tritone palmato" è molto simile al Tritone volgare, le due specie non si reperiscono nello stesso ambiente poiché competono per l'occupazione della medesima nicchia ecologica; Triturus alpestris a quote più alte; Triturus cristatus di taglia più grande (circa 19cm), esibisce aggressività conspecifica; Triturus italicus endemismo italiano, 7,5cm di lunghezza.

Euproctus platycephalus è una specie endemica della Sardegna, l'habitat è costituito da ruscelli montani dalle acque molto ossigenate, è privo di polmoni per cui respira solo attraverso cute e faringe, la carenza di ossigeno è un fattore limitante per l'espansione dell'areale; la popolazione ha scarsa consistenza numerica; gravemente minacciato di estinzione a causa dell'introduzione della trota, vorace predatore, a scopo di acquacoltura.

La famiglia Anfiumidae conta solo due specie endemiche degli USA sud-occidentali, acquatici, l'habitat sono i fiumi di grande portata, notturni, pedomorfici (conservano tratti dello stadio giovanile), lunghi circa un metro, corpo anguilliforme, arti estremamente ridotti, denti aguzzi, occhi privi di palpebre.

La famiglia Pletodontidae è la più diversificata, conta circa 300 specie conosciute; areale discontinuo (Nord America, America Centrale, Italia e Francia); morfologia e taglia molto variabili (dai 3cm ai 32cm); il carattere distintivo è il solco naso-labiale; mandibola fissa; assenza di polmoni; vivono in ambienti freschi che favoriscono respirazione cutanea; gregari (vivono in gruppo); spesso ovovivipari; forme cavernicole; in Italia e Francia sono rappresentati dai cosiddetti Geotritoni. Speleomantes ambrosii (Lanza, 1955) è endemico delle Alpi Apuane; misura mediamente 15cm di lunghezza; colorazione variabile (nero, grigio, marrone, oro); cavernicolo; occhi grandi e sporgenti specializzati all'oscurità; la lingua protrattile copre una distanza pari a più della metà del corpo; emancipati dall'ambiente acquatico, le uova vengono deposte in ambiente umido; la fecondazione indipendente dal singolo accoppiamento (la femmina conserva la spermateca anche per fecondazioni successive); dopo circa 8 mesi di sviluppo, l'uovo schiude in stadio già metamorfosato.
Proteidei
Il sottordine Proteidei, anche noto con il nome di Perennibranchiati, è contraddistinto da forme neoteniche che vivono in ambiente acquatico e che mantengono le branchie allo stadio adulto. Le branchie sono filamentose. L'areale discontinuo (distribuzione geografica disgiunta). Al sottordine fa capo un'unica famiglia omonima, Proteidae (Gray, 1825), che a sua volta include due soli generi:
- Necturus:
- del Nord America, l'habitat è rappresentato da acque superficiali e perenni, la pelle è pigmentata e gli occhi non del tutto regrediti, rappresentato dalla specie Necturus maculosus soprannominata "Cane d'acqua";
- Proteus:
- rappresentato dalla specie Proteus anguinus, endemica delle acque sotterranee del Carso e di una parte della ex-Jugoslavia, la pelle è apigmentata e gli occhi sono atrofizzati.

fotografia di Arne Hodalič.
Meanti
Il sottordine Meanti include l'unica famiglia Sirenidae (Gray, 1825) che si trova nella parte sud-orientale del Nord America e che a sua volta comprende due soli generi Siren (con due specie) e Pseudobranchus (con una sola specie). Neotenia obbligata (la somministrazione sperimentale di ormoni non induce la metamorfosi); acquatici; corpo anguilliforme; arti posteriori scomparsi, arti anteriori privi di cinto pelvico, crescita lenta e basso potenziale riproduttivo. In caso di siccità sono in grado di resistere immergendosi nel fango e racchiudendosi in un bozzolo di muco. Un rappresentate è Siren lacertina che misura circa 1m di lunghezza.


La morfologia dei membri appartenenti all'ordine dei Gimnofioni (detti anche Apodi) differisce notevolmente da quella generica degli Anfibi, in conseguenza dell'adattamento alla vita ipogea (sotterranea).
Per fenomeno di convergenza evolutiva, i Gimnofioni hanno evoluto alcuni tratti anatomici analoghi a quelli dei lombrichi (Anellidi): il corpo è estremamente allungato e a sezione circolare, dotato di pliche trasversali che conferiscono un aspetto analogo alla metameria dei lombrichi, la coda è assente per consentire alla cloaca di sboccare all'estremità posteriore.
Mostrano analogie anatomiche ed etologiche anche con i serpenti (Ofidi): i polmoni sono asimmetrici, la ventilazione si svolge nel solo polmone destro mentre il sinistro risulta atrofizzato; numerosi denti retrocurvi; aumento del numero di vertebre (alcune specie di Gimnofioni raggiungono il numero di 300 vertebre); cure parentali verso le uova, come i serpenti, certi Gimnofioni le proteggono cingendole con il proprio corpo fino al momento della schiusa.
Il maschio possiede un organo copulatore, perciò la fecondazione è interna. Spesso Ovipari, alcuni ovovivipari. Cranio di forma conica e fortemente ossificato. Il capo è provvisto di un organo di senso che consiste in un tentacolo ricco di terminazioni nervose che può essere retratto in un apposito alloggiamento situato sulla mascella. Gli occhi sono estremamente ridotti, privi di palpebre ma coperti da uno strato protettivo di pelle. Canale timpanico chiuso e non visibile all'esterno, l'udito è quasi del tutto assente. Mostrano un carattere esclusivo nell'ambito degli Anfibi: sono provvisti di scaglie, sebbene estremamente ridotte, che proteggono la pelle dall'attrito con il sottosuolo. Le dimensioni variano da 15cm a 130cm a seconda della specie. Sono carnivori, si nutrono prevalentemente di anellidi con cui condividono l'ambiente. L'habitat è rappresentato dall'ecotono ripario (biotopo che occupa la fascia che decorre parallelamente a contatto dei corsi d'acqua). Oltre 150 specie descritte, raggruppate in 5 famiglie, delle quali i Cecilidi è la più nota. Distribuiti in America centrale, Sud America, Africa centrale e Asia meridionale (compreso l'arcipelago indiano).


Ringraziamenti & Bibliografia
La presente dissertazione è ricavata dalla rielaborazione e dall'integrazione degli appunti e delle registrazioni audio di lezioni, nonché lucidi e dispense, relative a due distinti corsi universitari di zoologia; per cui i miei principali ringraziamenti vanno ai rispettivi docenti di tali corsi.
Sentiti ringraziamenti agli autori delle fotografie che mettono le proprie opere a libera disposizione di chiunque, e alle repository che le archiviano, tra cui principalmente Wikimedia Commons, ma anche ClipArtetc e Arthursclipart.
Grazie al Professor Pietro Omodeo per aver rinunciato ai diritti del suo testo rendendolo di pubblico dominio, e grazie all'ANISN di diffonderlo.
Grazie agli sviluppatori dell'applicazione grafica open source KolourPaint con cui ho personalmente prodotto le illustrazione che si trovano in questo ed in altri post del mio blog.
Uno speciale ringraziamento alla piattaforma blogger che ospita il mio blog e che mi regala la possibilità di esprimere la mia passione per la scrittura.
Ringrazio Roxana Minuz per l'attenta segnalazione dei refusi.
Ed infine grazie ai lettori del mio blog, perché senza di essi non sarei motivato a dare sfogo alla mia passione per la scrittura.